生物学、バイオテクノロジー、生態学の各分野に影響を与える画期的な開発において、東京理科大学付属地球生命科学研究所 (ELSI) の科学者たちは、次のような革新的なコンセプトを発表しました。 大域的制約原則。この新しい生物学的法則は、生命科学における長年の謎を説明します。 明らかに資源が豊富な状況であっても、なぜ生物は無限に成長しないのか。この発見は古典的なモデルの限界を超え、単細胞微生物から複雑な生態系に至るまで、生物学的成長の境界を理解するための統一された数学的に根拠のある枠組みを提供します。
この記事では、この発見を詳細に解き明かし、その理論的起源、実験による検証、および広範囲にわたる影響をたどります。基本的な生物学的法則を再検討し、刺激的な「段々樽」のメタファーを探求し、計算モデリングを検証します。 大腸菌、そしてこの原則が農業、医学、環境科学をどのように再構築できるかを考えてみましょう。
永続的なパズル: なぜ食料の増加は無限の成長を意味しないのか?
1 世紀以上にわたり、生物学者は生物システム全体にわたる一貫したパターンを観察してきました。 より多くの栄養素を追加すると成長が促進されますが、それはある程度までです。このしきい値を超えると、リソースを追加しても利益は減少します。として知られるこの現象は、 収穫逓減の法則 生物学では、ペトリ皿での細菌培養から肥沃な畑での作物の収量まで、あらゆるもので文書化されています。
簡単な実験室での実験を考えてみましょう: あなたは成長します 大腸菌 栄養のあるスープの中で。グルコース濃度が増加すると、細胞分裂が飽和点まで加速します。その後、砂糖を2倍にしても成長速度は2倍にはなりません。実際、それはほとんど変わりません。なぜ?
これは、これらの管理された環境での毒性や老廃物の蓄積によるものではありません。細胞は健康で、資源は豊富ですが、成長は停滞しています。何十年もの間、科学者たちはこれを「制限要因」によるものだと考えてきましたが、 機構 制限の背後にあるものはとらえどころのないままでした。それは 1 つのマスター ボトルネックでしたか?それとももっと複雑なものでしょうか?
古典の再訪: モノードとリービッヒの再検討
新しい発見を文脈で説明するには、まず伝統的な成長理論の 2 つの柱を理解する必要があります。
1. モノーの方程式 (1942)
フランスの生物学者ジャック モノーによって開発されたこのモデルは、微生物の増殖を次の関数として説明します。 単一の制限栄養素:
どこ:
- メートル = 比成長率
- メートルmax = 可能な最大成長率
- [S] = 基質(栄養素)濃度
- Ks= 半飽和定数
Monod のモデルは、制御された環境では非常にうまく機能しますが、次のことを前提としています。 たった1つの栄養素が成長を制御する 一度に。セルを外部供給に応答するブラック ボックスとして扱います。
2. リービッヒの最小の法則 (1840)
ドイツの化学者ユストゥス・フォン・リービッヒによって提案されたこの農業原則では、次のように述べられています。
「成長は、利用可能な総資源ではなく、最も希少な資源によって制限されます。」
高さの異なる板で作られた樽を想像してください。水位 (成長) は、最も短い板によって制限されます (栄養素が制限されます)。このモデルは直感的でありながら、 単一のボトルネックに焦点を当てる。
どちらのフレームワークも強力ですが、不完全です。彼らは説明に失敗している なぜ、たとえ「最短譜」が長くなったとしても(例えば、制限栄養素をさらに追加することによって)、成長は依然として無限に加速するわけではありません。何か別のことが人生を妨げているに違いありません。
グローバル制約原理: 限界のネットワーク
ELSIチームは次のように主張している。 人生は 1 つのボトルネックによって制限されるのではなく、相互依存する制約の動的なネットワークによって制限されます。 状況の変化に応じて変化します。彼らの洞察は、成長を外部供給のみへの反応としてではなく、 内部のバランスをとる行為 物理的、生化学的、空間的制限によって支配されます。
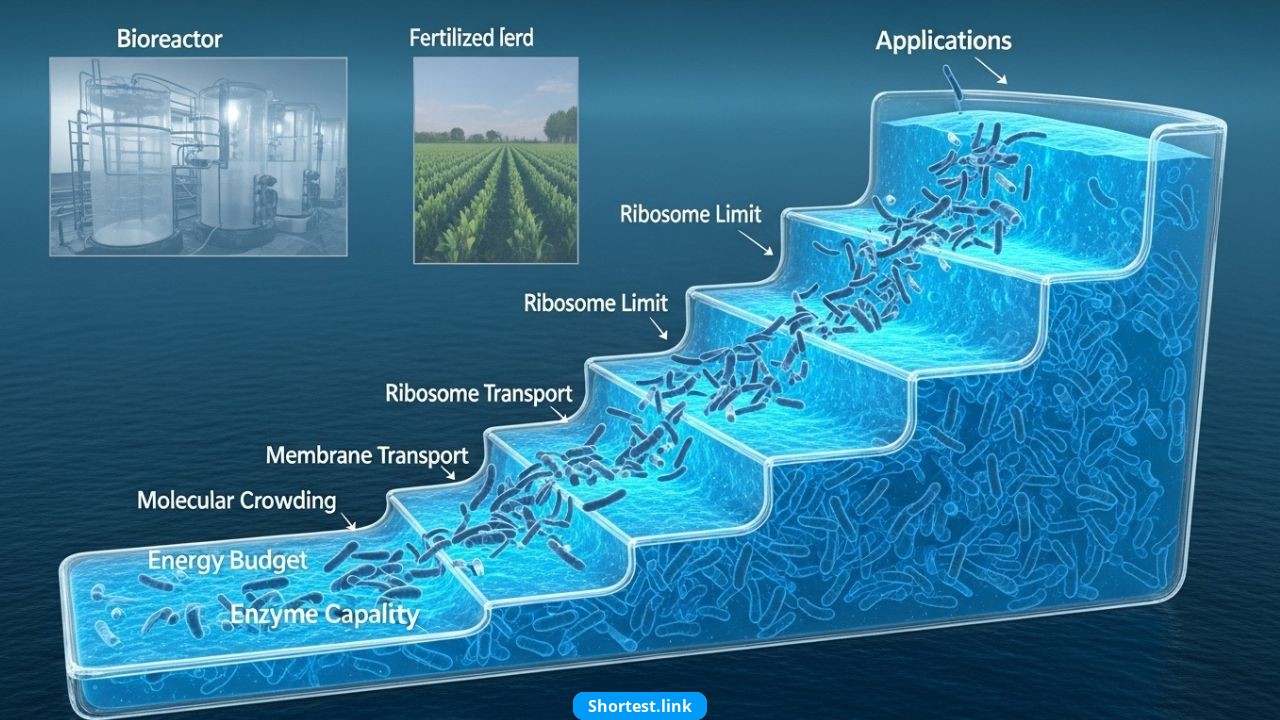
「テラスバレル」のメタファー
これを説明するために、研究者らは強力な新しい画像を導入しました。 テラスバレル。
譜表が配置された樽を想像してください。 ステップ—高さが均一ではありません。さらに「栄養水」を注ぐと、液体は上昇しますが、すぐに次の低い段階に達します。そのステップが新しい制約になります。さらに水を追加すると、さらに別のステップが発生し、それぞれが異なる内部制限を表します。
各「ステップ」は、異なる生物学的制約に対応します。
- 酵素の利用可能性:栄養素を処理するのに十分な触媒タンパク質がありません。
- リボソーム容量: タンパク質合成のための限られた機械。
- 膜輸送: 細胞膜は栄養素を十分な速さで取り込むことができません。
- 細胞質の混雑: 分子が密集しすぎて効率的な拡散ができなくなります。
- エネルギー配分のトレードオフ: メンテナンスに費やされたリソースは成長には使用できません。
栄養素が増加すると、1 つの制約が緩和され、代わりに別の制約が適用されます。したがって、 成長は徐々に鈍化する、たとえ豊富であっても。
計算による検証: モデリング 大腸菌 制約下
仮説を検証するために、ELSI チームは次のことを採用しました。 制約ベースの代謝モデリング、複数の同時制限の下で細胞プロセスをシミュレートするシステム生物学的アプローチ。
モデルの主要コンポーネント
研究者らは、ゲノムスケールのモデルを構築した。 大腸菌 統合されたもの:
| タンパク質の配分 | 細胞が酵素や構造タンパク質を合成する能力には限界があります。 |
| 分子の密集 | 細胞質の体積には限界があります。高分子が多すぎると、拡散と結合が妨げられます。 |
| 膜表面積 | 栄養素輸送体は膜に埋め込まれています。限られたスペースでは摂取が制限されます。 |
| エネルギー予算 | ATP は成長、維持、ストレス応答の間で分配される必要があります。 |
| 転写・翻訳能力 | リボソームと RNA ポリメラーゼは有限の資源です。 |
この複数の制約フレームワークを使用して、栄養濃度の勾配にわたる成長をシミュレートしました。
結果: 段階的な成長曲線
シミュレーションにより明らかになったのは、 非線形、漸近的な成長曲線 それは驚くべき精度で現実世界の実験データと一致しました。さらに重要なことは、モデルが示したことです。 相転移 制限要因の場合:
| 低い | 炭素源(例:グルコース) | グルコースを追加すると大幅に増加 |
| 中くらい | アミノ酸生合成酵素 | 中程度の増加 |
| 高い | リボソームの利用可能性 / タンパク質合成 | 弱い増加 |
| 非常に高い | 膜輸送能力 | ほぼゼロの増加 |
この進行は、「テラスバレル」のアイデアを裏付けています。 1 つの制約が克服されると、別の制約が現れます、暴走成長を防ぎます。
「それは細胞ではありません 選ぶ 「細胞の内部経済は、外部にどれだけの食物があるかに関係なく、物理的および生化学的な上限に達するということです。」と研究の筆頭著者であるタナカ・アカリ博士は説明する。
この発見が重要な理由: 専門分野を超えた影響
グローバル制約原則は単なる理論上の好奇心ではなく、複数の領域で変革をもたらす可能性を秘めています。
1. 生物学理論: 生命の統一された枠組み
初めて、成長制限は生物特有の癖としてではなく、 物理学と資源経済学に根ざした普遍的な原理。このモデルは、細菌、酵母、植物細胞、さらには動物組織にも同様に適用されます。
これにより、以下の間のギャップが埋められます。
- 微生物生理学
- 細胞生物物理学
- 進化のトレードオフ
- システム生物学
それは次のことを示唆しています すべての生命は「制約された風景」の中で活動します それが進化の軌跡を形作るのです。
2. バイオテクノロジー: 細胞工場の最適化
産業用バイオテクノロジーは、インスリン、バイオ燃料、酵素などを生産するために人工微生物に依存しています。しかし、栄養素の最適化にもかかわらず、収量は頭打ちになることがよくあります。
グローバル制約原則を使用すると、エンジニアは次のことが可能になります。
- 識別する どれの 制約とは、任意の条件で生産を制限することです
- 細胞リソースのリダイレクト(トランスポーターやリボソームの過剰発現など)
- ボトルネックの変化を予測した動的な供給戦略を設計する
例: バイオエタノール生産では、単に糖を追加するだけでなく、膜グルコーストランスポーターを同時発現させる可能性があります。 そして シャペロンタンパク質は混雑ストレスを軽減します。
3. 農業: よりスマートな施肥
農家は日常的に窒素、リン、カリウムを施用していますが、作物の収量はある時点を過ぎると収穫量が減少することがわかります。
新しい原則はその理由を次のように説明しています。 土壌の栄養素が植物の吸収能力を超えると、ボトルネックは次のように移行します。
- 根の表面積
- 光合成酵素濃度(例:ルビスコ)
- 水の輸送効率
これは、精密農業アプローチをサポートします。 入力を電流制限ステップに合わせて調整する、廃棄物と環境流出を削減します。
4. 生態学: 生態系の反応を予測する
水生生態系では、栄養汚染(富栄養化)により藻類の発生が発生しますが、これらは最終的には崩壊します。従来のモデルでは、タイミングと規模を予測するのが困難でした。
全体的な制約の枠組みは、ブルームがリンや窒素だけではなく、次のような要因によって制限されることを示唆しています。 光透過、CO₂拡散、またはウイルス溶解能力—出現する制約 後 初期の栄養飽和状態。
これにより、気候モデルが改善されます。 多層的な生物学的フィードバック 炭素と栄養素の循環予測に活用します。
微生物を超えて: 原理はスケールアップするのか?
ELSI チームは、グローバル制約原則が単一セルをはるかに超えて拡張される可能性があると慎重に提案しています。
多細胞生物の場合
人間の成長を考えてみましょう。たとえ豊富な食べ物があっても、子供たちは無限に成長するわけではありません。思春期が成長を止めるのは飢餓のためではなく、 ホルモンシグナル、骨プレート融合、エネルギー配分のトレードオフ 支配的な制約になります。
同様に、「制御されない増殖」と見なされることが多いがん細胞も、血液供給を超えたり (酸素制限)、免疫反応を引き起こしたり、DNA 損傷を蓄積したりするなど、依然として物理的な限界に従います。
生態系内
森は無限に高く成長するわけではありません。ある時点で、 木部を通る水の輸送、 風ストレス、 または 栄養素の循環速度 太陽光や二酸化炭素に関係なく、限界が生じます。
階段状の樽は、分子から惑星までスケールしているようです。
課題と今後の方向性
この原則は説得力がありますが、次のような新たな疑問を引き起こします。
- 複雑な生物におけるすべての制約を定量化できるでしょうか? ヒト肝細胞内の分子密集度を測定することは、肝細胞内の分子密集度を測定することよりもはるかに困難です。 大腸菌。
- 制約はどのように相互作用するのでしょうか? それらは相加的、相乗的、または階層的ですか?
- 進化は制約の状況を「再配線」できるでしょうか? 一部の極限環境微生物は、他の細胞を麻痺させるような条件下でも繁殖しますが、内部のボトルネックが変化したのでしょうか?
ELSI チームは現在、合成生物学者と協力して、 変更された制約プロファイルを使用してセルを設計する、成長限界をさらに押し上げることができるかどうかをテストします。
結論: 境界内の生活
グローバル制約原則の発見は、パラダイム シフトを示します。これは、一本の「アキレス腱」が人生を制限するという時代遅れの概念を置き換えるものです。 より豊かでダイナミックなビジョン: 人生はトレードオフの交響曲であり、機会と限界の間で絶えず交渉しています。
田中博士は次のように述べています。
「人生は、怠けたり非効率だからといって成長しないわけではありません。成長するのです」 物理学と化学が許す限り正確に。そして今、私たちはついにそれらの境界の地図を手に入れました。」
気候危機、食糧不安、バイオテクノロジー革命の時代において、普遍的な成長限界を理解することは学術的に奥深いだけでなく、持続可能な未来を築くためにも不可欠です。
要約表: グローバル制約原則の主要な概念
| 収穫逓減の法則 | 栄養素を増やしても成長が遅い | 説明が必要な観察された現象 |
| モノーの方程式 | 1つの栄養素によって成長が制限される | 便利だが単純化しすぎている |
| リービッヒの法則 | 希少な資源により成長が制限される | 細胞内部の制約を無視する |
| テラスバレル | 比喩: 複数の段階的な制約 | 動的に変化する制限を視覚化する |
| 制約の種類 | 酵素、リボソーム、膜、クラウディング、エネルギー | 特定の生物学的ボトルネックを特定します |
| 大腸菌モデリング | マルチ制約シミュレーションは実際のデータと一致します | 理論を実験的に検証します |
| 専門分野を超えた影響 | バイオテクノロジー、農業、生態学、医学 | 最適化と予測を可能にします |
| 普遍 | 微生物から生態系まで応用できる可能性がある | 生物学の基本法則を示唆する |
Leave a Reply